Molucelar evidence of a reed warbler x great reed warbler hybrid (Acrocephalus scirpaceus X A. arundinaceus) in Belgium
Bengt Hansson, Walter Roggeman en Gunter De Smet
Abstract
We report on the second case of reed warbler X great reed warbler hybrid (Acrocephalus scirpaceus and A. aruninaceus).
The bird was captured during a standardised ringing session in Belguim (Kinrooi) in autumn 1999, and fell between the parental species in all measurements.
Molecular analyses of two mincrosatellite loci verified that the bird was a female that had a reed warbler father and a great reed warbler mother.
Introduction, method and results
During a standarised ringing session at Kinrooi, Limburg, Belgium, on 20 August 1999, Guido Knoops captured a first calendar year Acrocephalus-warbler that could not be determined to spieces.
Detailed measurements were taken (Table 1) and some bodyfeathers were collected (W.R., G.D.S.).
In all measured characters the bird was larger than reed warbler (Acrocephalus scripaceus) and smaller than great reed warbler (A. arundinaceus; Table 1).
In 1995; Beier et al. (1996, 1997) observed a territorial male Acrocephalus-warbler in Germany that sang a song containing elements of the normal song of both great reed and reed warblers.
That individual was revealed as a reed X great reed hybrid: it was intermediate in morphology and had genetic material from both observation led us to suspect that the Belgian bird was also the result of hybridisation between a reed warbler and a great warbler.
Our molecular analyses verified this.
From one of the collected body-feathers, DNA was extracted by using lysis buffer and phenol/chloroform extraction (cf. Hansson et al. 2000). The DNA was analysed in three ways (analyses performed by B.H. at the Molecular Population Biology Laboratory, Lund University, Sweden). First, we determined the sex of the bird with a molecular method based on the absence or presence of the female-specific W-chromosome (Fridolfsson and Ellegren 1999). This analysis showed thtat the examined individual was a female. Secondly, we amplified two different microsatellite loci (Pocc1, Bensch et al. 1997; Aar8, Hansson et al. 2000). These loci were analysed because great reed and reed warblers have non-overlapping allele frequencies (Fig. 1; Hansson et al. 2000, B. Hansson, unpublished). The Belgian bird had genetic material from both species, i.e. it was a reed X great reed warbler hybrid (Fig. 1). Finnally, to reveal the mother-species of the hybrid, we sequenced a short fragment (350 bp) of the mitochondrial cytochrome b gene using the universal primers L14842 and H14149 (Kocher et al. 1989). The sequence almost perfectly machted the great reed warbler cytochrome b sequence that was published on GeneBank (99% similarity with AJ004784), but differed to a large extent from the reed warbler sequence (88% similarity with AJ004771; http://www.ncbi.nlm.nih.gov/entrez/query.fcgi ).
Table 1 Morphological data of great reed warblers (Acrocephalus arundinanceus), reed warblers (A. scirpaceus) and the female reed X great reed warbler hybrid examined at Kinrooi, Lumburg, Belgium, 20 August 1999. All data concern female birds and are given in mm. Data on great reed and reed warblers are from Cramp (1992).
| | Great reed warbler | Reed warbler | Hybrid (a) |
| Wing | 92,3 | 65,4 | 76,5 |
| Tail | 70,4 | 49,7 | 60 |
| Bill (to skull) | 22,7 | 16,8 | 20,6 |
| Bill (to nostril) | 12,7 | 9 | 10,3 |
| Tarsus | 28,6 | 22,8 | 26,2 |
(a) Additional measurements of the Belguim hybrid, Distance to longest primary (mm; p1=46.5, p2=1.5, p3=0, p4=3, p5=6, p6=8, p7=11, p8=13, p9=16, p10=18, s1=21, p1=1mm longer than longest primary covert; notch of p2=10mm. Wing formula: tip of p2 nearly equal to p7. Bill (to feathering) = 20.6mm. Bill width (at distal edge of nostril) = 4.4mm. Weight = 14.6g.
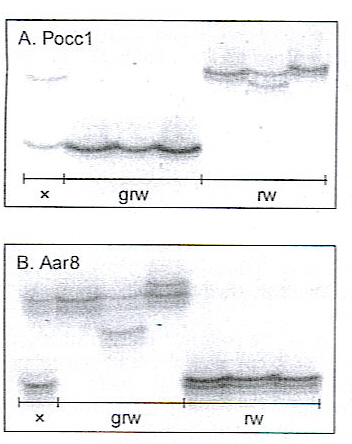
Fig. 1 Alleles at het microsatellite loci Pocc1 and Aar8 in the reed X great reed warbler (Acrocephalus scirpaceus X A. arundinaceus) hybrid (X), and great reed (grw) and reed warbler (rw) reference individuals.
Discussion
Our molecular analyses showed that the Acrophalus- warbler captured in Belgium was a female that had a reed warbler father and a great reed warbler mother. The same parental combination was found for te reed X great reed warbler hybrid examind bij Beier et al. (1997), although that individual was a male.
In addition to the two reed X great reed warbler hybrids that now hebe been observed in Germany and Belgium (Beier et al. 1996, 1997, this study), heterospecific matings and/or viable hybrids have occasionally been documented between other combinations of Acrocephalus spp., for example, between reed, marsh (A. palustris) and Blyth's reed (A. dumetorum) warblers (Lemaire 1997; Glutz von Blotzheim and Bauer 1985; Cramp 1992), and between great reed and clamorous reed (A. stentoreus) warblers (Hansson et al. 2004).
Thus, in the genus Acrocephalus both non-sister (reed and great reed warblers) and sister species (e.g. great reed and clamorous reed warblers) may hybridise (cf. Leisler et al. 1997; Helbig and Siebold 1999).
Avian hybridisation often occurs when two related species meet and one of them is rare (Grant and Grant 1992; Wirtz 1999; Randler 2002). Great reed and reed warblers breed sympatrically in reed lakes in Western Europe. In many of these lakes, great reed warblers are rare whereas reed warblers are common (Cramp 1992). Hence, our observation supports the hypothesis suggesting that hybridisation occus due to the relative scarcity of one of the parental species.
Bhansson
Departement of animal Ecology,lund University Sweden
W.Roggeman hoofd Belgisch ringwerk
G.De Smet
G.Knoops